
QUIEN DIJO QUE LA ANATOMIA ES ABURRIDA
martes, 22 de octubre de 2013
martes, 15 de octubre de 2013
lunes, 14 de octubre de 2013
jueves, 28 de febrero de 2013
CUERPO LUTEO
En caso de embarazo, el cuerpo lúteo crece hasta el comienzo del tercer mes de gestación, mantiene su función de producción hormonal y posteriormente va desapareciendo lentamente. Si el óvulo no es fecundado, y por tanto no es rescatado por la hCG embrionaria, el cuerpo lúteo desaparece en unos 10 o 12 días y la consiguiente disminución de la producción hormonal desencadena la menstruación. La insuficiencia de cuerpo lúteo, una patología que provoca un descenso de la concentración de progesterona en el plasma sanguíneo, es una de las principales causas de la Infertilidad femenina

sábado, 23 de febrero de 2013
SISTEMA ENDOCRINO
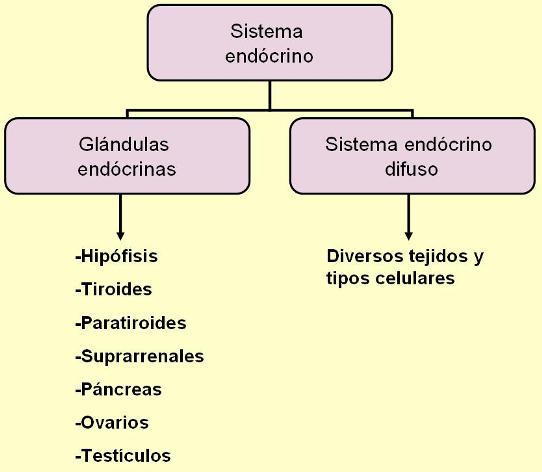
El sistema endócrino es el conjunto de las glándulas endócrinas o glándulas de secreción interna. Las glándulas endócrinas son órganos de origen epitelial especializados en la secreción, que carecen de conductos excretores. Sus secreciones, denominadas hormonas, difunden hacia la sangre y viajan por el torrente circulatorio.
Las hormonas son mensajeros químicos que producen efectos específicos en uno o varios tipos celulares u órganos a los que se llama células u órganos “blanco”. Los blancos de una hormona son aquellas estructuras que poseen un receptor específico para la misma.
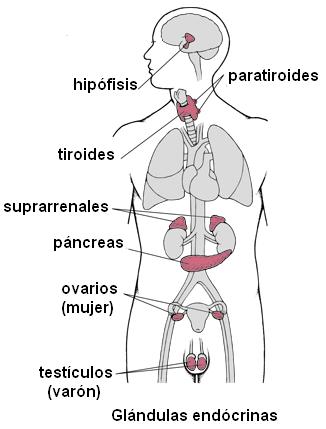
Las glándulas endócrinas son: la hipófisis o pituitaria, la tiroides, las paratiroides, las glándulas adrenales o suprarrenales, una porción del páncreas y las glándulas sexuales (ovarios y testículos).
Además de las glándulas endócrinas propiamente dichas, se incluyen dentro del sistema endócrino diversas células con la capacidad de liberar señales químicas que, ya sea a través del líquido intersticial o de la sangre, llegan hasta otras células blanco donde inducen una respuesta. Podría hablarse entonces de la existencia de un sistema endócrino difuso, formado por todas las células o tejidos corporales que, sin formar parte de una glándula endócrina, secretan mensajeros químicos hacia el medio interno.
Algunos componentes del sistema endócrino difuso son:
- Aparato digestivo: secreta un gran número de señales que controlan las funciones digestivas, como la colecistoquinina o el péptido VIP.
- Corazón: sintetiza el factor natriurético atrial, que controla la presión arterial.
- Riñón: produce eritropoyetina, que estimula la eritropoyesis, y renina, implicada en el control de la presión arterial.
- Endotelio: secreta prostaglandinas y muchas otras señales.
- Leucocitos: producen citoquinas, que modulan la respuesta inmune.
- Tejido adiposo: libera leptina, hormona que interviene en el control del apetito.
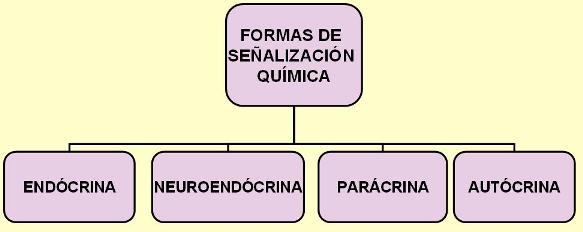
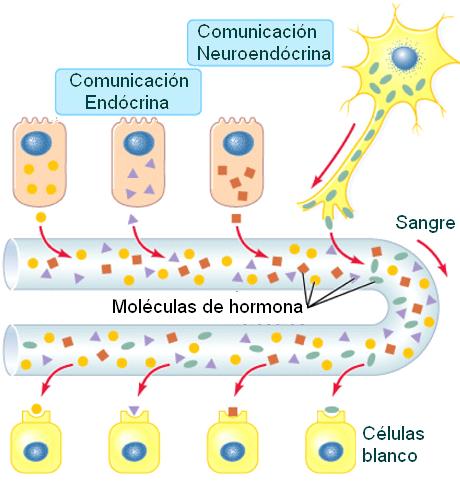
También ciertas neuronas pueden emitir señales químicas que se transportan por sangre. A las hormonas liberadas por células nerviosas se las denomina neurohormonas o secreciones neuroendócrinas.
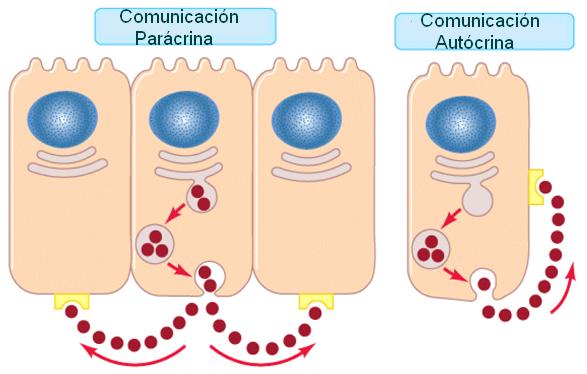
Las hormonas que difunden por el líquido intersticial y ejercen efectos sobre células vecinas reciben el nombre de secreciones parácrinas. En algunos casos, la célula endócrina es el blanco de su propia hormona. A este tipo de señalización se la denomina autócrina.
Las hormonas son mensajeros químicos que producen efectos específicos en uno o varios tipos celulares u órganos a los que se llama células u órganos “blanco”. Los blancos de una hormona son aquellas estructuras que poseen un receptor específico para la misma.
Las glándulas endócrinas son: la hipófisis o pituitaria, la tiroides, las paratiroides, las glándulas adrenales o suprarrenales, una porción del páncreas y las glándulas sexuales (ovarios y testículos).
Además de las glándulas endócrinas propiamente dichas, se incluyen dentro del sistema endócrino diversas células con la capacidad de liberar señales químicas que, ya sea a través del líquido intersticial o de la sangre, llegan hasta otras células blanco donde inducen una respuesta. Podría hablarse entonces de la existencia de un sistema endócrino difuso, formado por todas las células o tejidos corporales que, sin formar parte de una glándula endócrina, secretan mensajeros químicos hacia el medio interno.
Algunos componentes del sistema endócrino difuso son:
- Aparato digestivo: secreta un gran número de señales que controlan las funciones digestivas, como la colecistoquinina o el péptido VIP.
- Corazón: sintetiza el factor natriurético atrial, que controla la presión arterial.
- Riñón: produce eritropoyetina, que estimula la eritropoyesis, y renina, implicada en el control de la presión arterial.
- Endotelio: secreta prostaglandinas y muchas otras señales.
- Leucocitos: producen citoquinas, que modulan la respuesta inmune.
- Tejido adiposo: libera leptina, hormona que interviene en el control del apetito.
También ciertas neuronas pueden emitir señales químicas que se transportan por sangre. A las hormonas liberadas por células nerviosas se las denomina neurohormonas o secreciones neuroendócrinas.
Las hormonas que difunden por el líquido intersticial y ejercen efectos sobre células vecinas reciben el nombre de secreciones parácrinas. En algunos casos, la célula endócrina es el blanco de su propia hormona. A este tipo de señalización se la denomina autócrina.
Glándula hipófisis y eje hipotálamo-hipofisario Regresar
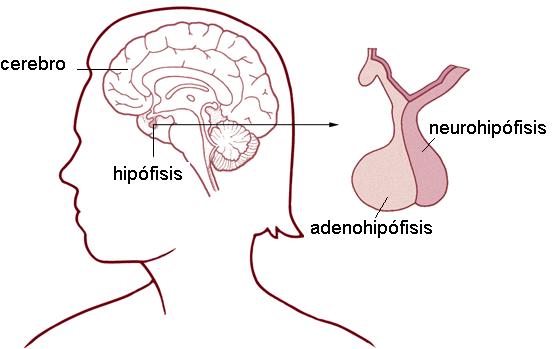
La glándula hipófisis o pituitaria tiene el tamaño de un guisante y se halla ubicada en el cráneo, apoyada sobre la silla turca del hueso esfenoides
La hipófisis está formada por dos lóbulos:
• Lóbulo anterior o adenohipófisis (adeno: glándula). Éste comprende las porciones tuberal, distal e intermedia.
• Lóbulo posterior o neurohipófisis
• Lóbulo anterior o adenohipófisis (adeno: glándula). Éste comprende las porciones tuberal, distal e intermedia.
• Lóbulo posterior o neurohipófisis
En el embrión, la adenohipófisis se origina a partir del epitelio de la cavidad oral y está formada por tejido glandular, mientras que la neurohipófisis surge como una excrecencia del diencéfalo (una parte del cerebro) y está formada por tejido nervioso. La neurohipófisis permanece conectada al hipotálamo, un núcleo gris del diencéfalo, a través del infundíbulo o tallo pituitario.
Además de la relación ontogénica (por su origen embrionario) y anatómica, el hipotálamo y la hipófisis mantienen una estrecha conexión funcional; las funciones de la hipófisis dependen directamente del hipotálamo y ambos forman una unidad fisiológica: el eje hipotálamo-hipofisario.
La relación funcional entre la hipófisis y el hipotálamo tiene dos aspectos:
La relación funcional entre la hipófisis y el hipotálamo tiene dos aspectos:
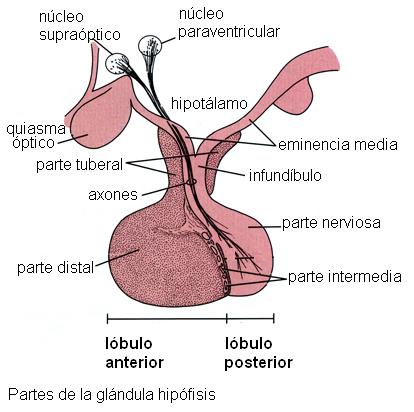
) La secreción de las hormonas sintetizadas en la adenohipófisis está bajo el control de factores liberadores o inhibidores producidos en el hipotálamo. Estos factores hipotalámicos son sintetizados en neuronas del hipotálamo y llegan hasta las células glandulares del lóbulo anterior mediante la circulación (comunicación neuroendócrina). La comunicación neuroendócrina existente entre el hipotálamo y la adenohipófisis depende del sistema porta hipofisario.
Un sistema porta es una red capilar interpuesta entre dos venas o dos arterias, a diferencia de los capilares comunes, que se interponen entre una arteria y una vena. Los capilares que drenan la sangre del hipotálamo reciben las neurosecreciones y luego se unen formando venas. Estas venas se capilarizan nuevamente al llegar al lóbulo anterior. La segunda red capilar (sistema porta) permite la difusión de los factores hipotalámicos hacia el espacio intersticial y desde allí hacia las células glandulares de la hipófisis anterior, a las cuales controlan. Esta conexión corta y directa entre el hipotálamo y la hipófisis favorece una respuesta rápida, impidiendo que la escasa cantidad de neurohormonas se diluya en la circulación general.
Un sistema porta es una red capilar interpuesta entre dos venas o dos arterias, a diferencia de los capilares comunes, que se interponen entre una arteria y una vena. Los capilares que drenan la sangre del hipotálamo reciben las neurosecreciones y luego se unen formando venas. Estas venas se capilarizan nuevamente al llegar al lóbulo anterior. La segunda red capilar (sistema porta) permite la difusión de los factores hipotalámicos hacia el espacio intersticial y desde allí hacia las células glandulares de la hipófisis anterior, a las cuales controlan. Esta conexión corta y directa entre el hipotálamo y la hipófisis favorece una respuesta rápida, impidiendo que la escasa cantidad de neurohormonas se diluya en la circulación general.
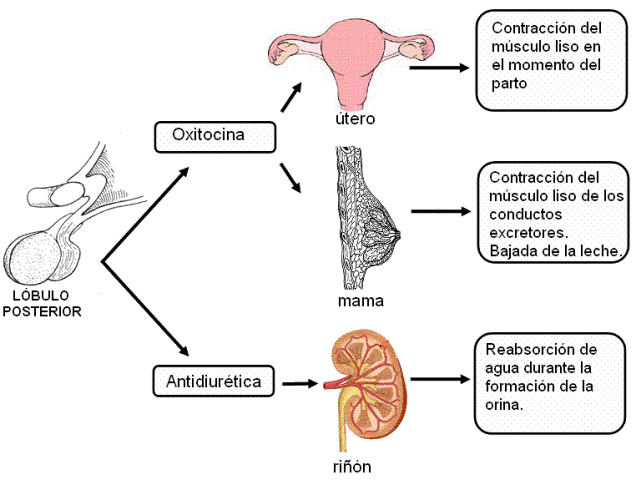
2) La neurohipófisis libera dos hormonas, la oxitocina y la hormona antidiurética (HAD). Estas hormonas son sintetizadas en los cuerpos de neuronas ubicadas en núcleos del hipotálamo (núcleos supraóptico y paraventricular). Una vez sintetizadas descienden por los axones de dichas neuronas, a través del infundíbulo. Los terminales axónicos se encuentran en el lóbulo posterior y desde allí las hormonas son liberadas a la circulación
Hormonas de la hipófisis Regresar
El lóbulo anterior de la hipófisis secreta hormonas tróficas, que actúan estimulando el desarrollo y la función de otros órganos. El lóbulo posterior secreta oxitocina y vasopresina, también llamada hormona antidiurética (HAD). Las hormonas y los efectos específicos sobre sus órganos blanco se resumen en el siguiente cuadro
Glándulas tiroides y paratiroides Regresar
La glándula tiroides es bilobulada y está ubicada por delante de la tráquea.
Las glándulas paratiroides son cuatro pequeñas glándulas (dos superiores y dos inferiores) situadas en la cara posterior de la tiroides
Las glándulas paratiroides son cuatro pequeñas glándulas (dos superiores y dos inferiores) situadas en la cara posterior de la tiroides
Glándulas suprarrenales Regresar
También llamadas adrenales, son dos glándulas ubicadas en los polos superiores de los riñones. Cada una posee una corteza y una médula, que tienen diferente origen embrionario y diferente función. La corteza está formada por tejido epitelial, en tanto la médula es de origen nervioso. Las secreciones de la médula adrenal son neurohormonas
Páncreas Regresar
El páncreas es una glándula mixta. El páncreas exócrino secreta enzimas digestivas que, mediante los conductos pancreáticos, se vuelcan en el intestino delgado.
El páncreas endócrino está conformado por grupos de células endócrinas distribuidos entre el tejido exócrino de la glándula. Las agrupaciones de células endócrinas se conocen como islotes de Langerhans.
En los islotes de Langerhans, distintos tipos celulares (alfa, beta, delta y F) se especializan en la síntesis de diferentes hormonas, como se detalla a continuación
El páncreas endócrino está conformado por grupos de células endócrinas distribuidos entre el tejido exócrino de la glándula. Las agrupaciones de células endócrinas se conocen como islotes de Langerhans.
En los islotes de Langerhans, distintos tipos celulares (alfa, beta, delta y F) se especializan en la síntesis de diferentes hormonas, como se detalla a continuación
Regulación del sistema endócrino Regresar
La secreción hormonal es regulada de forma que las concentraciones de hormonas en sangre sean las apropiadas para desencadenar la respuesta necesaria.
Las glándulas endócrinas son reguladas directamente por estímulos nerviosos procedentes de los nervios autónomos.
También son importantes los estímulos bioquímicos, como en el caso de la secreción de insulina, que es disparada por un aumento de la glucemia.
El hipotálamo y la hipófisis regulan la secreción en otras glándulas endócrinas. Tomemos por ejemplo el control en la secreción de hormonas tiroideas.
El hipotálamo secreta un factor liberador (TRH) que estimula a las células de la adenohipófisis especializadas en la síntesis de TSH. La TSH estimula a la glándula tiroides para que sintetice y secrete las hormonas tiroideas, T3 y T4. Pero cuando el nivel en sangre de T3 y T4 es el adecuado, la estimulación por parte del eje hipotálamo-hipofisario debe cesar. De lo contrario, se producirían síntomas por hiperfunción de la glándula.
La hipófisis y, en menor medida, el hipotálamo, detectan el nivel elevado de T3 y T4 en sangre. Ante esta señal, el hipotálamo disminuye la síntesis del factor liberador y la hipófisis frena la secreción de TSH. Así se logra mantener una concentración adecuada de T3 y T4. Este mecanismo de control recibe el nombre de retroalimentación negativa: el efecto (nivel de T3 y T4) suprime la causa (factor liberador y TSH).
Cuando los niveles de T3 y T4 descienden más allá de lo conveniente, el hipotálamo y la hipófisis reanudan sus secreciones y el ciclo se reinicia
Las glándulas endócrinas son reguladas directamente por estímulos nerviosos procedentes de los nervios autónomos.
También son importantes los estímulos bioquímicos, como en el caso de la secreción de insulina, que es disparada por un aumento de la glucemia.
El hipotálamo y la hipófisis regulan la secreción en otras glándulas endócrinas. Tomemos por ejemplo el control en la secreción de hormonas tiroideas.
El hipotálamo secreta un factor liberador (TRH) que estimula a las células de la adenohipófisis especializadas en la síntesis de TSH. La TSH estimula a la glándula tiroides para que sintetice y secrete las hormonas tiroideas, T3 y T4. Pero cuando el nivel en sangre de T3 y T4 es el adecuado, la estimulación por parte del eje hipotálamo-hipofisario debe cesar. De lo contrario, se producirían síntomas por hiperfunción de la glándula.
La hipófisis y, en menor medida, el hipotálamo, detectan el nivel elevado de T3 y T4 en sangre. Ante esta señal, el hipotálamo disminuye la síntesis del factor liberador y la hipófisis frena la secreción de TSH. Así se logra mantener una concentración adecuada de T3 y T4. Este mecanismo de control recibe el nombre de retroalimentación negativa: el efecto (nivel de T3 y T4) suprime la causa (factor liberador y TSH).
Cuando los niveles de T3 y T4 descienden más allá de lo conveniente, el hipotálamo y la hipófisis reanudan sus secreciones y el ciclo se reinicia
Definición Regresar
El sistema endócrino es el conjunto de las glándulas endócrinas o glándulas de secreción interna. Las glándulas endócrinas son órganos de origen epitelial especializados en la secreción, que carecen de conductos excretores. Sus secreciones, denominadas hormonas, difunden hacia la sangre y viajan por el torrente circulatorio.
Las hormonas son mensajeros químicos que producen efectos específicos en uno o varios tipos celulares u órganos a los que se llama células u órganos “blanco”. Los blancos de una hormona son aquellas estructuras que poseen un receptor específico para la misma.
Las glándulas endócrinas son: la hipófisis o pituitaria, la tiroides, las paratiroides, las glándulas adrenales o suprarrenales, una porción del páncreas y las glándulas sexuales (ovarios y testículos).
Además de las glándulas endócrinas propiamente dichas, se incluyen dentro del sistema endócrino diversas células con la capacidad de liberar señales químicas que, ya sea a través del líquido intersticial o de la sangre, llegan hasta otras células blanco donde inducen una respuesta. Podría hablarse entonces de la existencia de un sistema endócrino difuso, formado por todas las células o tejidos corporales que, sin formar parte de una glándula endócrina, secretan mensajeros químicos hacia el medio interno.
Algunos componentes del sistema endócrino difuso son:
- Aparato digestivo: secreta un gran número de señales que controlan las funciones digestivas, como la colecistoquinina o el péptido VIP.
- Corazón: sintetiza el factor natriurético atrial, que controla la presión arterial.
- Riñón: produce eritropoyetina, que estimula la eritropoyesis, y renina, implicada en el control de la presión arterial.
- Endotelio: secreta prostaglandinas y muchas otras señales.
- Leucocitos: producen citoquinas, que modulan la respuesta inmune.
- Tejido adiposo: libera leptina, hormona que interviene en el control del apetito.
También ciertas neuronas pueden emitir señales químicas que se transportan por sangre. A las hormonas liberadas por células nerviosas se las denomina neurohormonas o secreciones neuroendócrinas.
Las hormonas que difunden por el líquido intersticial y ejercen efectos sobre células vecinas reciben el nombre de secreciones parácrinas. En algunos casos, la célula endócrina es el blanco de su propia hormona. A este tipo de señalización se la denomina autócrina.
Las hormonas son mensajeros químicos que producen efectos específicos en uno o varios tipos celulares u órganos a los que se llama células u órganos “blanco”. Los blancos de una hormona son aquellas estructuras que poseen un receptor específico para la misma.
Las glándulas endócrinas son: la hipófisis o pituitaria, la tiroides, las paratiroides, las glándulas adrenales o suprarrenales, una porción del páncreas y las glándulas sexuales (ovarios y testículos).
Además de las glándulas endócrinas propiamente dichas, se incluyen dentro del sistema endócrino diversas células con la capacidad de liberar señales químicas que, ya sea a través del líquido intersticial o de la sangre, llegan hasta otras células blanco donde inducen una respuesta. Podría hablarse entonces de la existencia de un sistema endócrino difuso, formado por todas las células o tejidos corporales que, sin formar parte de una glándula endócrina, secretan mensajeros químicos hacia el medio interno.
Algunos componentes del sistema endócrino difuso son:
- Aparato digestivo: secreta un gran número de señales que controlan las funciones digestivas, como la colecistoquinina o el péptido VIP.
- Corazón: sintetiza el factor natriurético atrial, que controla la presión arterial.
- Riñón: produce eritropoyetina, que estimula la eritropoyesis, y renina, implicada en el control de la presión arterial.
- Endotelio: secreta prostaglandinas y muchas otras señales.
- Leucocitos: producen citoquinas, que modulan la respuesta inmune.
- Tejido adiposo: libera leptina, hormona que interviene en el control del apetito.
También ciertas neuronas pueden emitir señales químicas que se transportan por sangre. A las hormonas liberadas por células nerviosas se las denomina neurohormonas o secreciones neuroendócrinas.
Las hormonas que difunden por el líquido intersticial y ejercen efectos sobre células vecinas reciben el nombre de secreciones parácrinas. En algunos casos, la célula endócrina es el blanco de su propia hormona. A este tipo de señalización se la denomina autócrina.
 |
 |
 |
Glándula hipófisis y eje hipotálamo-hipofisario Regresar
La glándula hipófisis o pituitaria tiene el tamaño de un guisante y se halla ubicada en el cráneo, apoyada sobre la silla turca del hueso esfenoides.
 |
La hipófisis está formada por dos lóbulos:
• Lóbulo anterior o adenohipófisis (adeno: glándula). Éste comprende las porciones tuberal, distal e intermedia.
• Lóbulo posterior o neurohipófisis.
• Lóbulo anterior o adenohipófisis (adeno: glándula). Éste comprende las porciones tuberal, distal e intermedia.
• Lóbulo posterior o neurohipófisis.
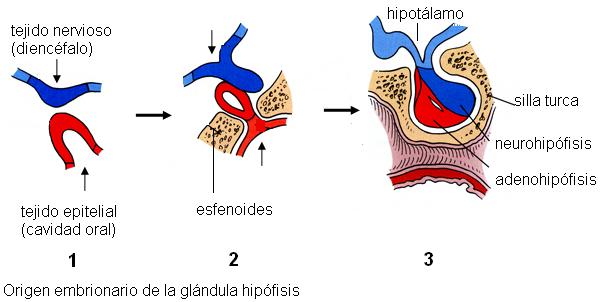 |
En el embrión, la adenohipófisis se origina a partir del epitelio de la cavidad oral y está formada por tejido glandular, mientras que la neurohipófisis surge como una excrecencia del diencéfalo (una parte del cerebro) y está formada por tejido nervioso. La neurohipófisis permanece conectada al hipotálamo, un núcleo gris del diencéfalo, a través del infundíbulo o tallo pituitario.
 |
Además de la relación ontogénica (por su origen embrionario) y anatómica, el hipotálamo y la hipófisis mantienen una estrecha conexión funcional; las funciones de la hipófisis dependen directamente del hipotálamo y ambos forman una unidad fisiológica: el eje hipotálamo-hipofisario.
La relación funcional entre la hipófisis y el hipotálamo tiene dos aspectos:
La relación funcional entre la hipófisis y el hipotálamo tiene dos aspectos:
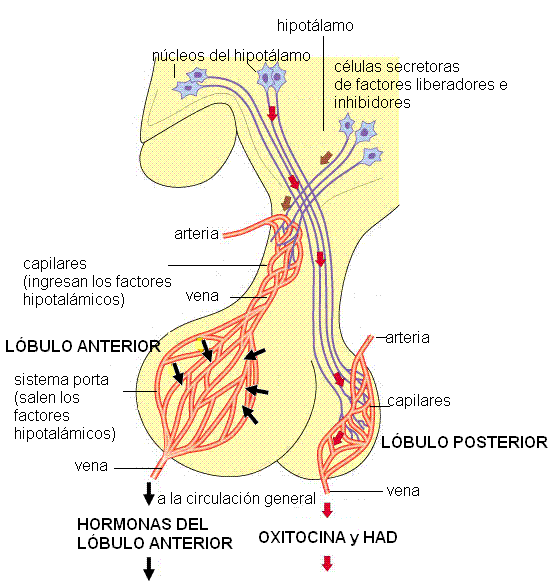 |
1) La secreción de las hormonas sintetizadas en la adenohipófisis está bajo el control de factores liberadores o inhibidores producidos en el hipotálamo. Estos factores hipotalámicos son sintetizados en neuronas del hipotálamo y llegan hasta las células glandulares del lóbulo anterior mediante la circulación (comunicación neuroendócrina). La comunicación neuroendócrina existente entre el hipotálamo y la adenohipófisis depende del sistema porta hipofisario.
Un sistema porta es una red capilar interpuesta entre dos venas o dos arterias, a diferencia de los capilares comunes, que se interponen entre una arteria y una vena. Los capilares que drenan la sangre del hipotálamo reciben las neurosecreciones y luego se unen formando venas. Estas venas se capilarizan nuevamente al llegar al lóbulo anterior. La segunda red capilar (sistema porta) permite la difusión de los factores hipotalámicos hacia el espacio intersticial y desde allí hacia las células glandulares de la hipófisis anterior, a las cuales controlan. Esta conexión corta y directa entre el hipotálamo y la hipófisis favorece una respuesta rápida, impidiendo que la escasa cantidad de neurohormonas se diluya en la circulación general.
Un sistema porta es una red capilar interpuesta entre dos venas o dos arterias, a diferencia de los capilares comunes, que se interponen entre una arteria y una vena. Los capilares que drenan la sangre del hipotálamo reciben las neurosecreciones y luego se unen formando venas. Estas venas se capilarizan nuevamente al llegar al lóbulo anterior. La segunda red capilar (sistema porta) permite la difusión de los factores hipotalámicos hacia el espacio intersticial y desde allí hacia las células glandulares de la hipófisis anterior, a las cuales controlan. Esta conexión corta y directa entre el hipotálamo y la hipófisis favorece una respuesta rápida, impidiendo que la escasa cantidad de neurohormonas se diluya en la circulación general.
2) La neurohipófisis libera dos hormonas, la oxitocina y la hormona antidiurética (HAD). Estas hormonas son sintetizadas en los cuerpos de neuronas ubicadas en núcleos del hipotálamo (núcleos supraóptico y paraventricular). Una vez sintetizadas descienden por los axones de dichas neuronas, a través del infundíbulo. Los terminales axónicos se encuentran en el lóbulo posterior y desde allí las hormonas son liberadas a la circulación.
Hormonas de la hipófisis Regresar
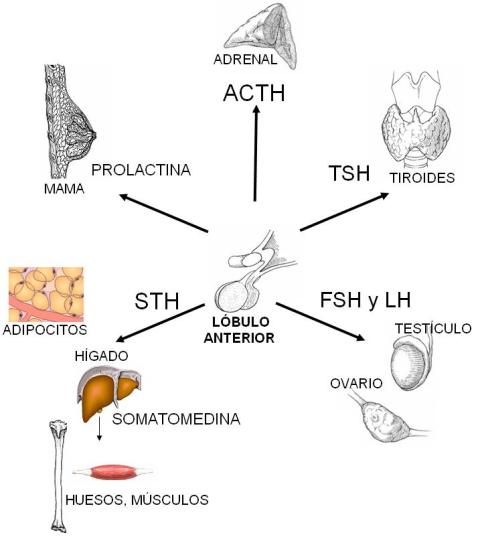
El lóbulo anterior de la hipófisis secreta hormonas tróficas, que actúan estimulando el desarrollo y la función de otros órganos. El lóbulo posterior secreta oxitocina y vasopresina, también llamada hormona antidiurética (HAD). Las hormonas y los efectos específicos sobre sus órganos blanco se resumen en el siguiente cuadro.
|
| ||
Somatotrofina (STH)
| Es la hormona del crecimiento. Estimula la captación de aminoácidos por las células, promoviendo la síntesis de proteínas. Estimula la secreción de somatomedinas (factores de crecimiento) en el hígado. Éstas promueven el crecimiento de distintos órganos y especialmente del cartílago, determinando el aumento en longitud de los huesos. En el tejido adiposo estimula la lipólisis. La liberación de ácidos grasos para la producción de energía produce un ahorro de proteínas, que se destinan al crecimiento. | ||
| Prolactina | Promueve el crecimiento y desarrollo de las glándulas mamarias durante el embarazo. Fomenta la lactancia después del parto. | ||
| Adrenocorticotrofina (ACTH) | Estimula la secreción de glucorticoides y aldosterona en la corteza de la glándula suprarrenal. | ||
| Tirotrofina (TSH) | Estimula el crecimiento de la glándula tiroides y la secreción de hormonas tiroideas. | ||
| Gonadotrofina: Hormona Foliculo-estimulante (FSH) | Estimula la gametogénesis, tanto en el varón como en la mujer. Regula el ciclo menstrual. | ||
| Gonadotrofina: Hormona Luteinizante (LH) | Estimula la producción de hormonas sexuales, tanto en el varón como en la mujer. Regula el ciclo menstrual. |
 |
Glándulas tiroides y paratiroides Regresar
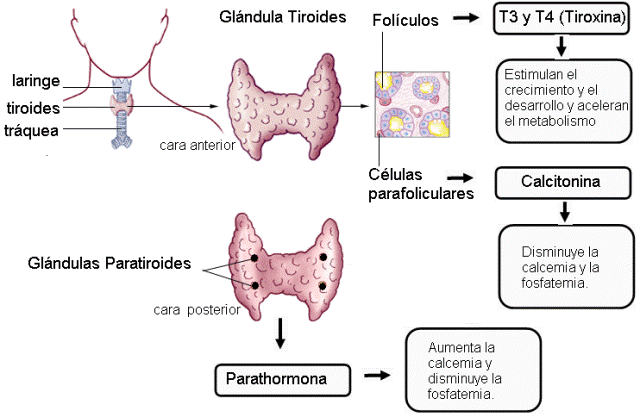
La glándula tiroides es bilobulada y está ubicada por delante de la tráquea.
Las glándulas paratiroides son cuatro pequeñas glándulas (dos superiores y dos inferiores) situadas en la cara posterior de la tiroides.
Las glándulas paratiroides son cuatro pequeñas glándulas (dos superiores y dos inferiores) situadas en la cara posterior de la tiroides.
Glándulas suprarrenales Regresar
También llamadas adrenales, son dos glándulas ubicadas en los polos superiores de los riñones. Cada una posee una corteza y una médula, que tienen diferente origen embrionario y diferente función. La corteza está formada por tejido epitelial, en tanto la médula es de origen nervioso. Las secreciones de la médula adrenal son neurohormonas.
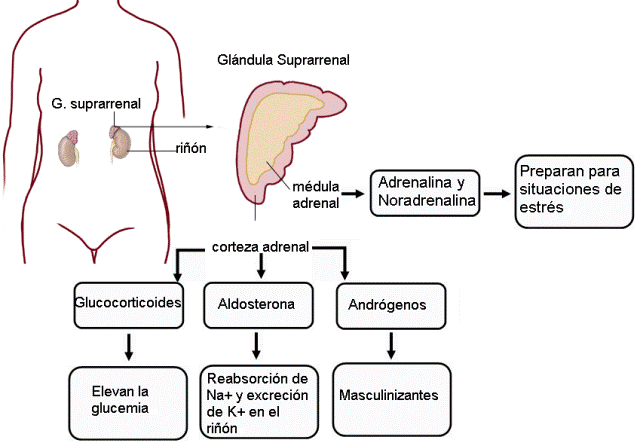 |
Páncreas Regresar
El páncreas es una glándula mixta. El páncreas exócrino secreta enzimas digestivas que, mediante los conductos pancreáticos, se vuelcan en el intestino delgado.
El páncreas endócrino está conformado por grupos de células endócrinas distribuidos entre el tejido exócrino de la glándula. Las agrupaciones de células endócrinas se conocen como islotes de Langerhans.
En los islotes de Langerhans, distintos tipos celulares (alfa, beta, delta y F) se especializan en la síntesis de diferentes hormonas, como se detalla a continuación.
El páncreas endócrino está conformado por grupos de células endócrinas distribuidos entre el tejido exócrino de la glándula. Las agrupaciones de células endócrinas se conocen como islotes de Langerhans.
En los islotes de Langerhans, distintos tipos celulares (alfa, beta, delta y F) se especializan en la síntesis de diferentes hormonas, como se detalla a continuación.
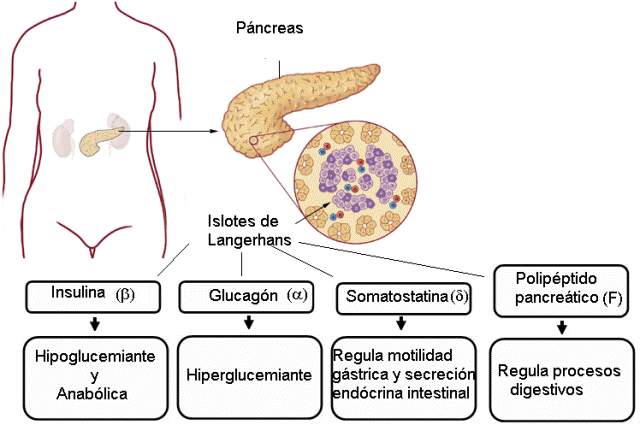
Regulación del sistema endócrino Regresar
La secreción hormonal es regulada de forma que las concentraciones de hormonas en sangre sean las apropiadas para desencadenar la respuesta necesaria.
Las glándulas endócrinas son reguladas directamente por estímulos nerviosos procedentes de los nervios autónomos.
También son importantes los estímulos bioquímicos, como en el caso de la secreción de insulina, que es disparada por un aumento de la glucemia.
El hipotálamo y la hipófisis regulan la secreción en otras glándulas endócrinas. Tomemos por ejemplo el control en la secreción de hormonas tiroideas.
El hipotálamo secreta un factor liberador (TRH) que estimula a las células de la adenohipófisis especializadas en la síntesis de TSH. La TSH estimula a la glándula tiroides para que sintetice y secrete las hormonas tiroideas, T3 y T4. Pero cuando el nivel en sangre de T3 y T4 es el adecuado, la estimulación por parte del eje hipotálamo-hipofisario debe cesar. De lo contrario, se producirían síntomas por hiperfunción de la glándula.
La hipófisis y, en menor medida, el hipotálamo, detectan el nivel elevado de T3 y T4 en sangre. Ante esta señal, el hipotálamo disminuye la síntesis del factor liberador y la hipófisis frena la secreción de TSH. Así se logra mantener una concentración adecuada de T3 y T4. Este mecanismo de control recibe el nombre de retroalimentación negativa: el efecto (nivel de T3 y T4) suprime la causa (factor liberador y TSH).
Cuando los niveles de T3 y T4 descienden más allá de lo conveniente, el hipotálamo y la hipófisis reanudan sus secreciones y el ciclo se reinicia.
Las glándulas endócrinas son reguladas directamente por estímulos nerviosos procedentes de los nervios autónomos.
También son importantes los estímulos bioquímicos, como en el caso de la secreción de insulina, que es disparada por un aumento de la glucemia.
El hipotálamo y la hipófisis regulan la secreción en otras glándulas endócrinas. Tomemos por ejemplo el control en la secreción de hormonas tiroideas.
El hipotálamo secreta un factor liberador (TRH) que estimula a las células de la adenohipófisis especializadas en la síntesis de TSH. La TSH estimula a la glándula tiroides para que sintetice y secrete las hormonas tiroideas, T3 y T4. Pero cuando el nivel en sangre de T3 y T4 es el adecuado, la estimulación por parte del eje hipotálamo-hipofisario debe cesar. De lo contrario, se producirían síntomas por hiperfunción de la glándula.
La hipófisis y, en menor medida, el hipotálamo, detectan el nivel elevado de T3 y T4 en sangre. Ante esta señal, el hipotálamo disminuye la síntesis del factor liberador y la hipófisis frena la secreción de TSH. Así se logra mantener una concentración adecuada de T3 y T4. Este mecanismo de control recibe el nombre de retroalimentación negativa: el efecto (nivel de T3 y T4) suprime la causa (factor liberador y TSH).
Cuando los niveles de T3 y T4 descienden más allá de lo conveniente, el hipotálamo y la hipófisis reanudan sus secreciones y el ciclo se reinicia.
Un tipo de regulación diferente se produce en el caso de la oxitocina. Durante el embarazo, la prolactina y otras hormonas estimulan el desarrollo de la mama, preparándola para la producción de leche. Sin embargo, la leche no baja hasta que se coloca el bebé al pecho y éste comienza a succionar. La eyección de la leche depende de la contracción del músculo liso de la glándula, estimulado por la oxitocina. La secreción de oxitocina es la respuesta de un reflejo nervioso disparado por el estímulo de la succión.
En este ejemplo se observa la integración de los controles nervioso y endócrino, y además un tipo de retroalimentación positiva.
Cuanto más se estimula el pezón, mayor es la secreción de oxitocina y mayor la cantidad de leche secretada. Por eso, las mujeres pueden amamantar por largos períodos, mientras mantengan la frecuencia de la lactancia. Si, en cambio, dejan de amamantar o disminuyen la frecuencia, el estímulo es cada vez menor y la secreción de leche empieza a disminuir hasta que cesa.
En este ejemplo se observa la integración de los controles nervioso y endócrino, y además un tipo de retroalimentación positiva.
Cuanto más se estimula el pezón, mayor es la secreción de oxitocina y mayor la cantidad de leche secretada. Por eso, las mujeres pueden amamantar por largos períodos, mientras mantengan la frecuencia de la lactancia. Si, en cambio, dejan de amamantar o disminuyen la frecuencia, el estímulo es cada vez menor y la secreción de leche empieza a disminuir hasta que cesa.
Composición química de las hormonas Regresar
Desde el punto de vista químico, las hormonas pueden agruparse en cuatro grandes clases:
1) Péptidos, proteínas, glucoproteínas: pueden ser cadenas muy cortas o de alto PM. Se sintetizan en el sistema de endomembranas y se almacenan en gránulos secretorios hasta la exocitosis. Incluyen a las hormonas de hipotálamo, hipófisis, páncreas y paratiroides.
Composición química de las hormonas Regresar
Desde el punto de vista químico, las hormonas pueden agruparse en cuatro grandes clases:
1) Péptidos, proteínas, glucoproteínas: pueden ser cadenas muy cortas o de alto PM. Se sintetizan en el sistema de endomembranas y se almacenan en gránulos secretorios hasta la exocitosis. Incluyen a las hormonas de hipotálamo, hipófisis, páncreas y paratiroides.
Composición química de las hormonas Regresar
Desde el punto de vista químico, las hormonas pueden agruparse en cuatro grandes clases:
1) Péptidos, proteínas, glucoproteínas: pueden ser cadenas muy cortas o de alto PM. Se sintetizan en el sistema de endomembranas y se almacenan en gránulos secretorios hasta la exocitosis. Incluyen a las hormonas de hipotálamo, hipófisis, páncreas y paratiroides.
Composición química de las hormonas Regresar
Desde el punto de vista químico, las hormonas pueden agruparse en cuatro grandes clases:
1) Péptidos, proteínas, glucoproteínas: pueden ser cadenas muy cortas o de alto PM. Se sintetizan en el sistema de endomembranas y se almacenan en gránulos secretorios hasta la exocitosis. Incluyen a las hormonas de hipotálamo, hipófisis, páncreas y paratiroides.
2) Derivados de aminoácidos: son pequeñas moléculas hidrosolubles. Se sintetizan en el citosol y luego se introducen en vesículas donde son reservadas. Por ejemplo: hormonas tiroideas.
3) Esteroides: derivan del colesterol. Son liposolubles. Se sintetizan en REL y mitocondrias. No se almacenan. Son las hormonas corticoadrenales y sexuales y la vitamina D.
4) Eicosanoides: derivan de un ácido graso (ácido araquidónico). Se sintetizan en la membrana plasmática y no se almacenan. Actúan como mensajeros locales (parácrinos). Son las prostaglandinas y los leucotrienos, producidos en casi todos los tipos celulares.
Mecanismo de acción de las hormonas en las células blanco Regresar
Las hormonas ejercen sus efectos a través de su unión al receptor. Los receptores de hormonas son proteínas. Cuando se unen la hormona y el receptor, este último experimenta un cambio conformacional. El receptor activado por la hormona inicia la vía de “transducción de la señal”. Ésta consiste en una cascada de modificaciones en la fisiología de la célula blanco que, directa o indirectamente, produce lo que llamamos la “respuesta” a la hormona.
Cada tipo de receptor dispara una vía de transducción particular. A su vez, esta vía es diferente según el tipo de célula. Dado que para una hormona pueden existir varios receptores distintos, esto explica la diversidad de respuestas inducidas por la misma hormona en los órganos blanco.
Hay dos grandes tipos de receptores hormonales: los que se encuentran insertos en la membrana plasmática y los intracelulares.
Los receptores de membrana reconocen a hormonas de naturaleza hidrofílica, por ejemplo a las hormonas proteicas. Este tipo de moléculas, poco afines a los lípidos, no puede atravesar la membrana celular y por lo tanto el receptor debe estar ubicado en la superficie de la célula blanco.
Las hormonas lipofílicas, como los esteroides, difunden fácilmente a través de las bicapas lipídicas. Estas hormonas traspasan la membrana plasmática y se unen a un receptor intracelular.
Cada tipo de receptor dispara una vía de transducción particular. A su vez, esta vía es diferente según el tipo de célula. Dado que para una hormona pueden existir varios receptores distintos, esto explica la diversidad de respuestas inducidas por la misma hormona en los órganos blanco.
Hay dos grandes tipos de receptores hormonales: los que se encuentran insertos en la membrana plasmática y los intracelulares.
Los receptores de membrana reconocen a hormonas de naturaleza hidrofílica, por ejemplo a las hormonas proteicas. Este tipo de moléculas, poco afines a los lípidos, no puede atravesar la membrana celular y por lo tanto el receptor debe estar ubicado en la superficie de la célula blanco.
Las hormonas lipofílicas, como los esteroides, difunden fácilmente a través de las bicapas lipídicas. Estas hormonas traspasan la membrana plasmática y se unen a un receptor intracelular.
Una vez formado el complejo hormona-receptor se desencadena el efecto fisiológico en la célula blanco.
a) En algunos casos, los receptores de membrana tienen una actividad enzimática que es “encendida” por la llegada de la hormona. Estos receptores catalizan la fosforilación (adición de grupos fosfato) de proteínas celulares, que a su vez se activan o se inactivan (según el caso) al ser fosforiladas. De esta forma se modifica la actividad celular.
b) Otros receptores de membrana actúan acoplados a una proteína G. Las proteínas G (de las que existen varios tipos) son las encargadas de inhibir o activar enzimas que fabrican “segundos mensajeros”. Los segundos mensajeros son como los “voceros” de la hormona (el primer mensajero) en el medio intracelular. La respuesta de la célula blanco depende del control que los segundos mensajeros ejercen sobre ciertas proteínas celulares. Un nucleótido derivado del ATP, el AMP cíclico (AMPc) y el calcio actúan frecuentemente como segundos mensajer
a) En algunos casos, los receptores de membrana tienen una actividad enzimática que es “encendida” por la llegada de la hormona. Estos receptores catalizan la fosforilación (adición de grupos fosfato) de proteínas celulares, que a su vez se activan o se inactivan (según el caso) al ser fosforiladas. De esta forma se modifica la actividad celular.
b) Otros receptores de membrana actúan acoplados a una proteína G. Las proteínas G (de las que existen varios tipos) son las encargadas de inhibir o activar enzimas que fabrican “segundos mensajeros”. Los segundos mensajeros son como los “voceros” de la hormona (el primer mensajero) en el medio intracelular. La respuesta de la célula blanco depende del control que los segundos mensajeros ejercen sobre ciertas proteínas celulares. Un nucleótido derivado del ATP, el AMP cíclico (AMPc) y el calcio actúan frecuentemente como segundos mensajer
Una vez formado el complejo hormona-receptor se desencadena el efecto fisiológico en la célula blanco.
a) En algunos casos, los receptores de membrana tienen una actividad enzimática que es “encendida” por la llegada de la hormona. Estos receptores catalizan la fosforilación (adición de grupos fosfato) de proteínas celulares, que a su vez se activan o se inactivan (según el caso) al ser fosforiladas. De esta forma se modifica la actividad celular.
b) Otros receptores de membrana actúan acoplados a una proteína G. Las proteínas G (de las que existen varios tipos) son las encargadas de inhibir o activar enzimas que fabrican “segundos mensajeros”. Los segundos mensajeros son como los “voceros” de la hormona (el primer mensajero) en el medio intracelular. La respuesta de la célula blanco depende del control que los segundos mensajeros ejercen sobre ciertas proteínas celulares. Un nucleótido derivado del ATP, el AMP cíclico (AMPc) y el calcio actúan frecuentemente como segundos mensajer
a) En algunos casos, los receptores de membrana tienen una actividad enzimática que es “encendida” por la llegada de la hormona. Estos receptores catalizan la fosforilación (adición de grupos fosfato) de proteínas celulares, que a su vez se activan o se inactivan (según el caso) al ser fosforiladas. De esta forma se modifica la actividad celular.
b) Otros receptores de membrana actúan acoplados a una proteína G. Las proteínas G (de las que existen varios tipos) son las encargadas de inhibir o activar enzimas que fabrican “segundos mensajeros”. Los segundos mensajeros son como los “voceros” de la hormona (el primer mensajero) en el medio intracelular. La respuesta de la célula blanco depende del control que los segundos mensajeros ejercen sobre ciertas proteínas celulares. Un nucleótido derivado del ATP, el AMP cíclico (AMPc) y el calcio actúan frecuentemente como segundos mensajer
Comparación de los sistemas endócrino y nervioso Regresar
El sistema nervioso y el endócrino trabajan estrechamente ligados. Existen muchos mecanismos por los cuales uno afecta la función del otro y en algunos casos sus funciones se solapan.
Los dos sistemas tienen características comunes. Ambos actúan por intermedio de mensajeros químicos, los neurotransmisores y las hormonas, aunque éstos viajan de manera diferente.
Tanto los neurotransmisores como las hormonas inducen efectos en células diana donde se localizan receptores específicos.
Las respuestas endócrinas son relativamente lentas, pero prolongadas. Las respuestas nerviosas son muy rápidas y también más fugaces.
El sistema nervioso permite respuestas a estímulos externos e internos y gobierna la relación con el ambiente. El sistema endócrino también está sometido a la influencia del ambiente, pero de manera indirecta, por intermedio del sistema nervioso. El sistema endócrino ejerce fundamentalmente el control del medio interno. Regula el metabolismo, la presión arterial, el crecimiento y el desarrollo, la reproducción y ciertos aspectos de la conducta.
Los dos sistemas tienen características comunes. Ambos actúan por intermedio de mensajeros químicos, los neurotransmisores y las hormonas, aunque éstos viajan de manera diferente.
Tanto los neurotransmisores como las hormonas inducen efectos en células diana donde se localizan receptores específicos.
Las respuestas endócrinas son relativamente lentas, pero prolongadas. Las respuestas nerviosas son muy rápidas y también más fugaces.
El sistema nervioso permite respuestas a estímulos externos e internos y gobierna la relación con el ambiente. El sistema endócrino también está sometido a la influencia del ambiente, pero de manera indirecta, por intermedio del sistema nervioso. El sistema endócrino ejerce fundamentalmente el control del medio interno. Regula el metabolismo, la presión arterial, el crecimiento y el desarrollo, la reproducción y ciertos aspectos de la conducta.
viernes, 1 de febrero de 2013
COXAL FEMUR PATELA
El miembro inferior se conecta al sistema esquelético axial a través del cíngulo pélvico formado por los dos hu
miembro inferior se cnoecta al sistema esquelético axial a través del cíngulo pélvico formado por los dos huesos coxales. El miembro inferior puede dividirse en tres segmentos: muslo, pierna y pie. El pie, a su vez, está formado por los huesos del tarso, metatarso y las falanges u ortejos.En el ángulo anterosuperior se ubica la espina iliaca anterosuperior
En el ángulo posterosuperior se ubica la espina iliaca posterosuperior
En el ángulo anteromedial se ubica el ángulo del pubis
En el ángulo posteroinferior se ubica la tuberosidad isquiática
El borde superior corresponde a la cresta ilíaca, lugar de inserción muscular abdominal lateral. El borde posterior presenta la incisura innominada, la espina ilíiaca posteroinferior, las incisuras isquiáticas mayor y menor, separadas por la espina isquiática. El borde anterior posee la incisura innominada, espina ilíaca anteroinferior. El margen inferior posee la rama isquiopubiana.
En su cara lateral se encuentra la cara articular para la cabeza femoral, el acetábulo, que es el lugar de la fusión de los tres huesos. El acetábulo en su profundidad presenta una superficie rugosa no articular conocida como fosa acetabular. Rodeando esta fosa, se encuentra la faceta lunata o semilunar. El acetábulo se encuentra limitado por la ceja o reborde acetabular que presenta tres incisuras; isquipubiana o acetabular, iliopubiana e ilioisquiática. El coxal presenta tres superficies articulares: el acetábulo, para la cabeza femoral; la cara sacropélvica que articula con el sacro (cara auricular y tuberosidad ilíaca); y, finalmente, la cara sinfisiaria del pubis que articula con el pubis contralateral.
Ilion
El ilion posee una ubicación superior en el coxal. Posee un cuerpo, que forma parte del acetábulo; y un ala. El ala posee dos caras: lateral, de orientación infero-lateral; y medial, de orientación supero-medial. La cara lateral o cara glútea presenta las líneas glúteas para la inserción de los músculos glúteos. La cara medial o fosa ilíaca sirve para la sujeción de las vísceras e inserción del músculo ilíaco. La cara sacropélvica presenta la cara auricular para articular con el sacro y la tuberosidad ilíaca para la inserción del ligamento. Presenta en su cara medial la línea arqueada para la distribución de fuerzas.
Isquión
Posee una ubicación posteroinferior en el coxal. Se divide en cuerpo y rama. El cuerpo forma dos quintos de la pared del acetábulo y se une con el ilion y el pubis. El isquión se une con la rama del pubis y forma con el cuerpo de este la incisura acetabular, limitando el foramen obturado que casi esta totalmente cerrado por la membrana obturatriz en el ser vivo. Dorsalmente se observa la espina isquiática que separa las dos incisuras isquiáticas, mayor y menor. La incisura isquiática menor formada por el isquion. El isquion posee una rugosidad denominada tuberosidad isquiática que corresponde a la región dorso-caudal del ramo del isquion.
Ilion
El ilion posee una ubicación superior en el coxal. Posee un cuerpo, que forma parte del acetábulo; y un ala. El ala posee dos caras: lateral, de orientación infero-lateral; y medial, de orientación supero-medial. La cara lateral o cara glútea presenta las líneas glúteas para la inserción de los músculos glúteos. La cara medial o fosa ilíaca sirve para la sujeción de las vísceras e inserción del músculo ilíaco. La cara sacropélvica presenta la cara auricular para articular con el sacro y la tuberosidad ilíaca para la inserción del ligamento. Presenta en su cara medial la línea arqueada para la distribución de fuerzas.
Isquión
Posee una ubicación posteroinferior en el coxal. Se divide en cuerpo y rama. El cuerpo forma dos quintos de la pared del acetábulo y se une con el ilion y el pubis. El isquión se une con la rama del pubis y forma con el cuerpo de este la incisura acetabular, limitando el foramen obturado que casi esta totalmente cerrado por la membrana obturatriz en el ser vivo. Dorsalmente se observa la espina isquiática que separa las dos incisuras isquiáticas, mayor y menor. La incisura isquiática menor formad por el isquion. El isquion posee una rugosidad denominada tuberosidad isquiática que corresponde a la región dorso-caudal del ramo del isquion.
Ilion
El ilion posee una ubicación superior en el coxal. Posee un cuerpo, que forma parte del acetábulo; y un ala. El ala posee dos caras: lateral, de orientación infero-lateral; y medial, de orientación supero-medial. La cara lateral o cara glútea presenta las líneas glúteas para la inserción de los músculos glúteos. La cara medial o fosa ilíaca sirve para la sujeción de las vísceras e inserción del músculo ilíaco. La cara sacropélvica presenta la cara auricular para articular con el sacro y la tuberosidad ilíaca para la inserción del ligamento. Presenta en su cara medial la línea arqueada para la distribución de fuerzas.
Isquión
Posee una ubicación posteroinferior en el coxal. Se divide en cuerpo y rama. El cuerpo forma dos quintos de la pared del acetábulo y se une con el ilion y el pubis. El isquión se une con la rama del pubis y forma con el cuerpo de este la incisura acetabular, limitando el foramen obturado que casi esta totalmente cerrado por la membrana obturatriz en el ser vivo. Dorsalmente se observa la espina isquiática que separa las dos incisuras isquiáticas, mayor y menor. La incisura isquiática menor formada por el isquion. El isquion posee una rugosidad denominada tuberosidad isquiática que corresponde a la región dorso-caudal del ramo del isquion.


martes, 29 de enero de 2013
H I P O F I S I S
La hipófisis o glándula pituitaria es una glándula endocrina que segrega hormonas encargadas de regular la homeostasis incluyendo las hormonas trópicas que regulan la función de otras glándulas del sistema endocrino, dependiendo en parte del hipotálamo el cual a su vez regula la secreción de algunas hormonas. Es una glándula compleja que se aloja en un espacio óseo llamado silla turca del hueso esfenoides, situada en la base del cráneo, en la fosa cerebral media, que conecta con el hipotálamo a través del tallo pituitario o tallo hipofisario. Tiene forma ovalada con un diámetro anteroposterior de 8 mm, trasversal de 12 mm y 6 mm en sentido vertical, en promedio pesa en el hombre adulto 500 miligramos, en la mujer 600 mg y en las que han tenido varios partos, hasta 700 mg.

I N D I C E
- 1 Partes
- 2 Adenohipófisis
- 3 Neurohipófisis
- 4 Síndrome de Cushing
- 5 Regulación hipotalámica
- 6 Patología
- 7 Etimología de la palabra
- 8 Imágenes
- 9 Notas
- 10 Enlaces externos

- Lóbulo anterior o adenohipófisis: procede embriológicamente de un esbozo faríngeo (bolsa de Rathke) y es responsable de la secreción de numerosas hormonas (ver más adelante).
- Hipófisis media o pars intermedia: produce dos polipéptidos llamados melanotropinas u hormonas estimulantes de los melanocitos, que inducen el aumento de la síntesis de melanina de las células de la piel.
- Lóbulo posterior o neurohipófisis: procedente de la evaginación del piso del tercer ventrículo del diencéfalo, al cual se le conoce con el nombre de infundíbulo, queda unido a través del tallo hipofisario; almacena a las hormonas ADH y oxitocina secretadas por las fibras amielínicas de los núcleos supraópticos y paraventriculares de las neuronas del hipotálamo.
A D E N O H I P O F I S I S
La adenohipófisis segrega muchas hormonas de las cuales seis son relevantes para la función fisiológica adecuada del organismo, las cuales son segregadas por 5 tipos de células diferentes. Estas células son de origen epitelial y como muchas glándulas endocrinas, están organizadas en lagunas rodeadas de capilares sinusoides a los cuales se vierte su secreción hormonal. Los tipos de células se clasificaban antes de acuerdo a su tinción, y eran acidófilas, basófilas y cromófobas (o que no se tiñen), sin embargo, esta clasificación no aportaba información significativa sobre su actividad secretora, por lo que en la actualidad se usa una clasificación de acuerdo con técnicas de inmunohistoquímica para sus productos de secreción, y se han podido identificar 5 tipos celulares:
- Células somatótropas que segregan GH (acidófila).
- Células lactotropas, o mamótropas que segregan PRL (acidófila).
- Células corticótropas que segregan ACTH (basófila).
- Células gonadótropas que segregan las gonadotropinas LH, y FSH (basófila).
- Células tirotropas que secretan la TSH (basófila).
Las células cromófobas son en realidad células desgastadas y pueden haber sido cualquiera de las cinco anteriores.
HORMONA DE LA ADENOHIPOFISIS
- Hormona del crecimiento o somatotropina (GH). Estimula la síntesis proteica, e induce la captación de glucosa por parte del músculo y los adipocitos, además induce la gluconeogénesis por lo que aumenta la glucemia; su efecto más importante es quizás que promueve el crecimiento de todos los tejidos y los huesos en conjunto con las somatomedinas. Por lo que un déficit de esta hormona causa enanismo y un aumento (ocasionado por un tumor acidófilo) ocasiona gigantismo en niños, y acromegalia en adultos, (consecuencia del previo cierre de los discos epifisiarios).
- Prolactina (PRL) u hormona luteotrópica. Estimula el desarrollo de los acinos mamarios y estimula la traducción de los genes para las proteínas de la leche.
- Hormona estimulante del tiroides (TSH) o tirotropina. Estimula la producción de hormonas por parte del tiroides
- Hormona estimulante de la corteza suprarrenal (ACTH) o corticotropina. Estimula la producción de hormonas por parte de las glándulas suprarrenales
- Hormona luteinizante (LH). Estimulan la producción de hormonas por parte de las gónadas y la ovulación.
- Hormona estimulante del folículo (FSH). Complementa la función estimulante de las gónadas provocada por la (LH).
la LH y la FSH se denominan gonadotropinas, ya que regulan la función de las gónadas.
N E U R O H I P O F I S I S
La neurohipófisis tiene un origen embriológico diferente al del resto de la hipófisis, mediante un crecimiento hacia abajo del hipotálamo, por lo que tiene funciones diferentes. Se suele dividir a su vez en tres partes: eminencia media, infundibulo y pars nervosa, de las cuales la última es la más funcional. Las células de la neurohipófisis se conocen como pituicitos y no son más que células gliales de sostén. Por tanto, la neurohipófisis no es en realidad una glándula secretora ya que se limita a almacenar los productos de secreción del hipotálamo. En efecto, los axoplasmas de las neuronas de los núcleos hipotalamicos supraóptico y paraventricular secretan la ADH y la oxitocina respectivamente, que se almacenan en las vesículas de los axones que de él llegan a la neurohipófisis; dichas vesículas se liberan cerca del plexo primario hipofisiario en respuesta impulsos eléctricos por parte del hipotálamo.
- Hormona antidiurética (ADH) o vasopresina. Se secreta en estímulo a una disminución del volumen plasmático y como consecuencia de la disminución en la presión arterial que esto ocasiona, y su secreción aumenta la reabsorción de agua desde los túbulos colectores renales por medio de la translocación a la membrana de la acuaporina II; también provoca una fuerte vasoconstricción por lo que también es llamada vasopresina.
- Oxitocina. Estimula la contracción de las células mioepiteliales de las glándulas mamarias lo que causa la eyección de leche por parte de la mama, y se estimula por la succión, transmitiendo señales al hipotálamo (retroalimentación) para que secrete más oxitocina. Causa contracciones del músculo liso del útero en el orgasmo y también los típicos espasmos de la etapa final del parto.
SINDROME DE CUSHING
principios del siglo XX, el neurocirujano Harvey Cushing (EE.UU., 1869-1939) comenzó a estudiar la glándula pituitaria, observando los efectos de un mal funcionamiento. Descubrió que una excrecion excesiva de adrenocorticotropina (ACTH) alteraba el metabolismo y el crecimiento, y le dio el nombre de síndrome de Cushing. Esta enfermedad, provoca debilidad en los miembros y fragilidad de los huesos. Cushing describió la pituitaria como "directora de la orquesta endocrinica", pero hoy se sabe que el verdadero director es el hipotálamo. El Cushing es un síndrome que afecta a varios sistemas y órganos, se caracteriza por una hipersecreción de cortisol (generalmente debido a una hiperplasia de la hipófisis). Los síntomas del Cushing son:
- Cara redonda, rubicunda y congestiva, lo que se denomina "cara en luna llena".
- Acúmulo de grasas en el cuello y nuca, lo que se conoce como cuello de búfalo.
- Obesidad central (abdomen sobresaliente pero extremidades delgadas).
- Estrías violáceas en abdomen, muslos y mamas.
- Dolores de espalda.
- Amenorrea.
- Aumento de vello púbico en mujeres
REGULACION HIPOTALAMICA
La hipófisis y el hipotálamo están conectados por un sistema capilar denominado sistema portal, el cual proviene de la arteria carótida interna y del polígono de Willis e irriga primero al hipotálamo formando el plexo capilar primario, que drena en los vasos porta hipofisiarios que a su vez forman el plexo capilar hipofisiario.
La importancia de este sistema es que transporta las hormonas liberadoras o hipofisiotrópicas que secreta el hipotálamo con fines reguladores de la secreción adenohipofisiaria. Estas hormonas son:- Somatoliberina (GHRH). Estimula la secreción de GH por parte de la hipófisis.
- Corticoliberina (ARH). Estimula la secreción de ACTH por parte de la hipófisis.
- Tiroliberina (TRH). Estimula la secreción de TSH por parte de la hipófisis.
- Gonadoliberina (LHRH). Estimula la secreción de LH y FSH por parte de la hipófisis.
- Hormona inhibidora de la GH (GHIH) o somatostatina. Inhibe la secreción de la GH por parte de la hipófisis.
El proceso se realiza en el momento en que el sistema nervioso central recibe un estímulo, el hipotálamo recibe parte de ese estímulo y actúa sobre la hipófisis, a su vez, el hipotálamo secreta las respectivas hormonas en la adenohipófisis o libera las de la neurohipófisis; estas se incorporan a la circulación, viajan por medio de la sangre y son captados por receptores específicos ubicados en los órganos diana, un ejemplo es la captación de la TSH por parte de los lóbulos tiroideos de la glándula tiroides.
En ese momento el órgano diana, que en todo caso es cualquiera de las glándulas endocrinas comienzan a secretar sus propias hormonas, con lo que se envía un estímulo al sistema nervioso, específicamente al hipotálamo, o directamente a la hipófisis con lo cual se contrarresta el estímulo inicial.
P A T O L O G I A
Algunos trastornos asociados a la hipófisis incluyen:| Condición | Etiología | Hormona |
| Acromegalia | sobreproducción | hormona de crecimiento |
| Gigantismo | sobreproducción | hormona de crecimiento |
| Deficiencia de la hormona de crecimiento | baja producción | hormona de crecimiento |
| Síndrome de secreción inadecuada de la hormona antidiurética | sobreproducción | vasopresina |
| Diabetes insípida | baja producción | vasopresina |
| Síndrome de Sheehan | baja producción | cualquier hormona hipofisaria |
| Adenoma hipofisario | sobreproducción | cualquier hormona hipofisaria |
| Hipopituitarismo | baja producción | cualquier hormona hipofisaria |
Un tumor en los remanentes de la bolsa de Rathke puede ocasionar un craneofaringioma, el cual comprime a la glándula hipófisis.
ETIMOLOGIA DE LA PALABRA
El término «hipófisis» proviene del griego hipo (‘debajo’) y fisis (‘crecer’).[3]
Aristóteles creía que el moco nasal (llamado pītuīta en latín)]se generaba en el cerebro, y salía por la nariz.
En 1543, Vesalio escribió lo mismo: que el moco nasal procedía de esa glándula en el cerebro. De allí el nombre «pituitaria»: glándula generadora de moco. En el siglo siguiente (XVII) se deshizo el error (el moco no provenía del cerebro, sino del interior de la misma nariz), y la anatomía ha preferido renombrar a esa glándula «hipófisis». Desde 1723, en latín científico, se documenta el nombre «pituitaria» o «membrana pituitaria» (‘mucosa generadora de moco’) para la mucosa nasal o mucosa olfatoria.
MEMBRANA PLASMATICA
La membrana plasmática es una bicapa lipídica que delimita todas las células. Es una estructura laminada formada por fosfolípidos, glicolípidos y proteínas que rodea, limita, da forma y contribuye a mantener el equilibrio entre el interior (medio intracelular) y el exterior (medio extracelular) de las células. La membrana plasmática regula la entrada y salida de muchas sustancias entre el citoplasma y el medio extracelular. Es similar a las membranas que delimitan los orgánulos de células eucariotas.
Está compuesta por dos láminas que sirven de "contenedor" para el citosol y los distintos compartimentos internos de la célula, así como también otorga protección mecánica. Está formada principalmente por fosfolípidos (fosfatidiletanolamina y fosfatidilcolina), colesterol, glúcidos y proteínas (integrales y periféricas).Diagrama del orden de los lípidos anfipáticos para formar una bicapa lipídica. Las cabezas polares (de color amarillento) separan las colas hidrofóbicas (de color gris) del medio citosólico y extracelular.
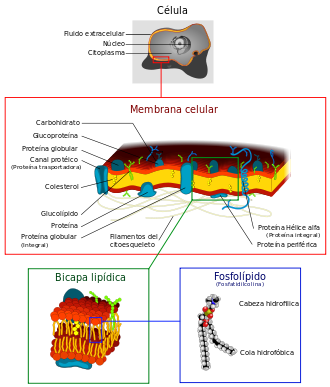

La principal característica de esta barrera es su permeabilidad selectiva, lo que le permite seleccionar las moléculas que deben entrar y salir de la célula. De esta forma se mantiene estable el medio intracelular, regulando el paso de agua, iones y metabolitos, a la vez que mantiene el potencial electroquímico (haciendo que el medio interno esté cargado negativamente). La membrana plasmática es capaz de recibir señales que permiten el ingreso de partículas a su interior.
Cuando una molécula de gran tamaño atraviesa o es expulsada de la célula y se invagina parte de la membrana plasmática para recubrirlas cuando están en el interior ocurren respectivamente los procesos de endocitosis y exocitosis.
Tiene un grosor aproximado de 7,5 nm y no es visible al microscopio óptico pero sí al microscopio electrónico, donde se pueden observar dos capas oscuras laterales y una central más clara. En las células procariotas y en las eucariotas osmótrofas como plantas y hongos, se sitúa bajo otra capa, denominada pared celular.
.jpg)
Esquema de una membrana celular. Según el modelo del Mosaico Fluido, las proteínas (en rojo y naranja) serían como "icebergs" que navegarían en lípidos (en azul). Nótese además que las cadenas de oligosacáridos (en verde) se hallan siempre en la cara externa, pero no en la interna.
I N D I C E
I N D I C E
Antiguamente se creía que la membrana plasmática era un conjunto estático formado por la sucesión de capas proteínas-lípidos-lípidos-proteínas. Hoy en día se concibe como una estructura dinámica cuyo modelo se conoce como "mosaico fluido", término acuñado por S. J. Singer y G. L. Nicolson en 1972. Esta estructura general -modelo unitario- se presenta también en todo el sistema de endomembranas (membranas de los diversos orgánulos del interior de la célula), como retículo endoplasmático, aparato de Golgi y envoltura nuclear, y los de otros orgánulos, como las mitocondrias y los plastos, que proceden de endosimbiosis.
La composición química de la membrana plasmática varía entre células dependiendo de la función o del tejido en la que se encuentren, pero se puede estudiar de forma general. La membrana plasmática está compuesta por una doble capa de fosfolípidos, por proteínas unidas no covalentemente a esa bicapa, y glúcidos unidos covalentemente a los lípidos o a las proteínas. Las moléculas más numerosas son las de lípidos, ya que se calcula que por cada 50 lípidos hay una proteína. Sin embargo, las proteínas, debido a su mayor tamaño, representan aproximadamente el 50% de la masa de la membrana.
B I C A P A L I P I D I C A
El orden de las llamadas cabezas hidrofílicas y las colas hidrofóbicas de la bicapa lipídica impide que solutos polares, como aminoácidos, ácidos nucleicos, carbohidratos, proteínas e iones, difundan a través de la membrana, pero generalmente permite la difusión pasiva de las moléculas hidrofóbicas. Esto permite a la célula controlar el movimiento de estas sustancias vía complejos de proteína transmembranal tales como poros y caminos, que permiten el paso de iones específicos como el sodio y el potasio.
Las dos capas de moléculas fosfolípidas forman un "sándwich" con las colas de ácido graso dispuestos hacia el centro de la membrana plasmática y las cabezas de fosfolípidos hacia los medios acuosos que se encuentran dentro y fuera de la célulaCOMPONENTES LIPIDICOS
El 98% de los lípidos presentes en las membranas celulares son anfipáticos, es decir que presentan un extremo hidrófilo (que tiene afinidad e interacciona con el agua) y un extremo hidrofóbico (que repele el agua). Los más abundantes son los fosfoglicéridos (fosfolípidos) y los esfingolípidos, que se encuentran en todas las células; le siguen los glucolípidos, así como esteroides (sobre todo colesterol). Estos últimos no existen o son escasos en las membranas plasmáticas de las células procariotas. Existen también grasas neutras, que son lípidos no anfipáticos, pero sólo representan un 2% del total de lípidos de membrana.

Diagrama del orden de los lípidos anfipáticos para formar una bicapa lipídica. Las cabezas polares (de color amarillento) separan las colas hidrofóbicas (de color gris) del medio citosólico y extracelular.

Diagrama del orden de los lípidos anfipáticos para formar una bicapa lipídica. Las cabezas polares (de color amarillento) separan las colas hidrofóbicas (de color gris) del medio citosólico y extracelular.
- Fosfoglicéridos. Tienen una molécula de glicerol con la que se esterifica un ácido fosfórico y dos ácidos grasos de cadena larga; los principales fosfoglicéridos de membrana son la fosfatidiletanolamina o cefalina, la fosfatidilcolina o lecitina, el fosfatidilinositol y la fosfatidilserina.
- Esfingolípidos. Son lípidos de membrana constituidos por ceramida (esfingosina + ácido graso); solo la familia de la esfingomielina posee fósforo; el resto poseen glúcidos y se denominan por ello glucoesfingolípidos o, simplemente glucolípidos. Los cerebrósidos poseen principalmente glucosa, galactosa y sus derivados (como N-acetilglucosamina y N-acetilgalactosamina). Los gangliósidos contienen una o más unidades de ácido N-acetilneuramínico (ácido siálico).
- Colesterol. El colesterol representa un 23% de los lípidos de membrana. Sus moléculas son pequeñas y más anfipáticas en comparación con otros lípidos. Se dispone con el grupo hidroxilo hacia el exterior de la célula (ya que ese hidroxilo interactúa con el agua). El colesterol es un factor importante en la fluidez y permeabilidad de la membrana ya que ocupa los huecos dejados por otras moléculas. A mayor cantidad de colesterol, menos permeable y fluida es la membrana. Se ha postulado que los lípidos de membrana se podrían encontrar en dos formas: como un líquido bidimensional, y de una forma más estructurada, en particular cuando están unidos a algunas proteínas formando las llamadas balsas lipídicas. Se cree que el colesterol podría tener un papel importante en la organización de estas últimas. Su función en la membrana plasmática es evitar que se adhieran las colas de ácido graso de la bicapa, mejorando la fluidez de la membrana. En las membranas de las células vegetales son más abundantes los fitoesteroles.
COMPONENTE PROTEICOS
El porcentaje de proteínas oscila entre un 20% en la vaina de mielina de las neuronas y un 70% en la membrana interna mitocondrial;[1] el 80% son intrínsecas, mientras que el 20% restantes son extrínsecas. Las proteínas son responsables de las funciones dinámicas de la membrana, por lo que cada membrana tienen una dotación muy específica de proteínas; las membranas intracelulares tienen una elevada proporción de proteínas debido al elevado número de actividades enzimáticas que albergan. En la membrana las proteínas desempeña diversas funciones: transportadoras, conectoras (conectan la membrana con la matriz extracelular o con el interior), receptoras (encargadas del reconocimiento celular y adhesión) y enzimas.
Las proteínas de la membrana plasmática se pueden clasificar según cómo se dispongan en la bicapa lipídica:
- Proteínas integrales. Embebidas en la bicapa lipídica, atraviesan la membrana una o varias veces, asomando por una o las dos caras (proteínas transmembrana); o bien mediante enlaces covalentes con un lípido o un glúcido de la membrana. Su aislamiento requiere la ruptura de la bicapa.
- Proteínas periféricas. A un lado u otro de la bicapa lipídica, pueden estar unidas débilmente por enlaces no covalentes. Fácilmente separables de la bicapa, sin provocar su ruptura.
- Proteína de membrana fijada a lípidos. Se localiza fuera de la bicapa lípidica, ya sea en la superficie extracelular o intracelular, conectada a los lípidos mediante enlaces covalentes.
En el componente proteico reside la mayor parte de la funcionalidad de la membrana; las diferentes proteínas realizan funciones específicas:
- Proteínas estructurales o de anclaje: estas proteínas hacen de "eslabón clave" uniéndose al citoesqueleto y la matriz extracelular.
- Proteínas receptoras: que se encargan de la recepción y transducción de señales químicas.
- Proteínas de transporte: mantienen un gradiente electroquímico mediante el transporte de membrana de diversos iones.
- Estas a su vez pueden ser:
- Proteínas transportadoras: Son enzimas con centros de reacción que sufren cambios conformacionales.
- Proteínas de canal: Dejan un canal hidrofílico por donde pasan los iones.
COMPONENTES GLUCIDOS
Están en la membrana unidos covalentemente a las proteínas o a los lípidos. Pueden ser polisacáridos u oligosacáridos. Se encuentran en el exterior de la membrana formando el glicocalix. Representan el 8% del peso seco de la membrana plasmática. Sus principales funciones son dar soporte a la membrana y el reconocimiento celular (colaboran en la identificación de las señales químicas de la célula).
F U N C I O N E S
- La función principal de la membrana plasmática es mantener el medio interno separado del externo. Esto es posible gracias a la naturaleza aislante en medio acuoso de la bicapa fosfolipídica y a las funciones de transporte que desempeñan las proteínas. La combinación de transporte activo y transporte pasivo hacen de la membrana endoplasmática una barrera selectiva que permite a la célula diferenciarse del medio.
- Permite a la célula dividir en secciones las distintas orgánelas y así proteger las reacciones químicas que ocurren en cada uno.
- Crea una barrera selectivamente permeable en donde solo entran o salen las sustancias estrictamente necesarias.
- Transporta sustancias de un lugar de la membrana a otro, ejemplo, acumulando sustancias en lugares específicos de la célula que le puedan servir para su metabolismo.
- Percibe y reacciona ante estímulos provocados por sustancias externas (ligandos).
- Mide las interacciones que ocurren entre células internas y externas.
P E R M E A B I L I D A D
La permeabilidad de las membranas es la facilidad de las moléculas para atravesarla. Esto depende principalmente de la carga eléctrica y, en menor medida, de la masa molar de la molécula. Moléculas pequeñas o con carga eléctrica neutra pasan la membrana más fácilmente que elementos cargados eléctricamente y moléculas grandes. Además, la membrana es selectiva, lo que significa que permite la entrada de unas moléculas y restringe la de otras. La permeabilidad depende de los siguientes factores:
- Solubilidad en los lípidos: Las sustancias que se disuelven en los lípidos (moléculas hidrófobas, no polares) penetran con facilidad en la membrana dado que está compuesta en su mayor parte por fosfolípidos.
- Tamaño: la mas grande parte de las moléculas de gran tamaño no pasan a través de la membrana. Sólo un pequeño número de moléculas polares de pequeño tamaño pueden atravesar la capa de fosfolípidos.
- Carga: Las moléculas cargadas y los iones no pueden pasar, en condiciones normales, a través de la membrana. Sin embargo, algunas sustancias cargadas pueden pasar por los canales proteicos o con la ayuda de una proteína transportadora.
- Canales: algunas proteínas forman canales llenos de agua por donde pueden pasar sustancias polares o cargadas eléctricamente que no atraviesan la capa de fosfolípidos.
- Transportadoras: otras proteínas se unen a la sustancia de un lado de la membrana y la llevan al otro lado donde la liberan.
La expresión membrana celular se usa con dos significados diferentes:
- Membrana plasmática, descrita en el presente artículo, es la membrana que siempre envuelve al citoplasma de las células. Aunque este uso siempre fue ilegítimo, está extraordinariamente extendido, sobre todo en los textos de habla inglesa (cell membrane).
- Pared celular, también llamada membrana de secreción, es una cubierta más o menos resistente que cubre a todas o la mayoría de las células de las plantas, los hongos y los protistas pluricelulares.
Durante siglo y medio (c.1800-c.1950) la investigación de las células se basó sólo en la observación mediante microscopía óptica. Ésta no puede, por razones físicas relacionadas con la longitud de onda de la luz, detectar estructuras de menos de 0,25 µm (micrómetros). Se llamó membrana celular al límite de la célula cuando éste era distinguible, y éste sigue siendo el único uso legítimo de la expresión. En la mayor parte de los casos lo que se observaba era un recubrimiento, más o menos flexible, hecho de polisacáridos, de proteínas o de polímeros mixtos, al que se llama también pared celular. Ésta es precisamente la expresión que debe preferirse para eludir la ambigüedad.
A principios del siglo XX, investigaciones experimentales de la fisiólogia celular condujeron a postular la existencia, en todas las células, de una membrana invisible, a la que se llamó membrana plasmática o citoplasmática, y que debía estar compuesta esencialmente de lípidos. Ésta representaba la envoltura del protoplasma, la parte fisiológicamente activada de la célula. Con el uso del microscopio electrónico, pudo observarse por fin la membrana plasmática, cuyo espesor típico es de sólo 0,0075 µm (109 Å).
Suscribirse a:
Entradas (Atom)